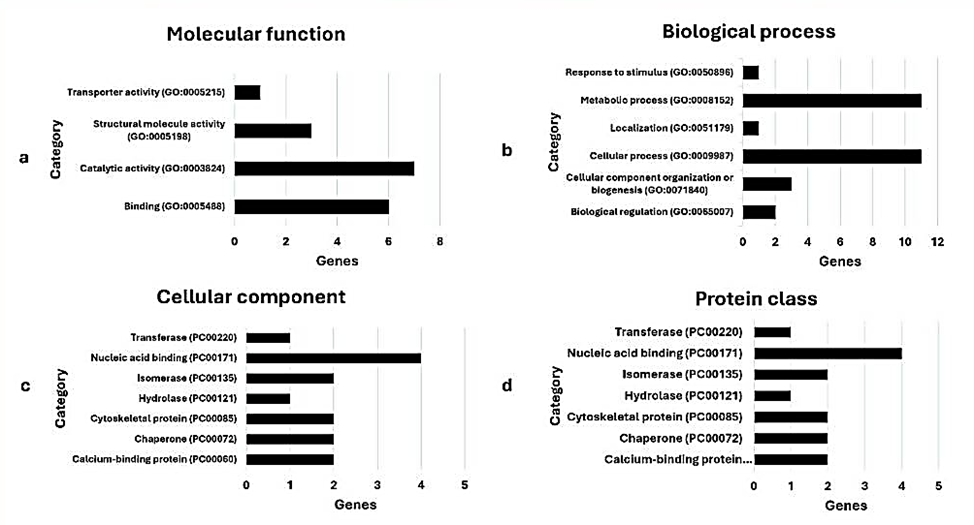
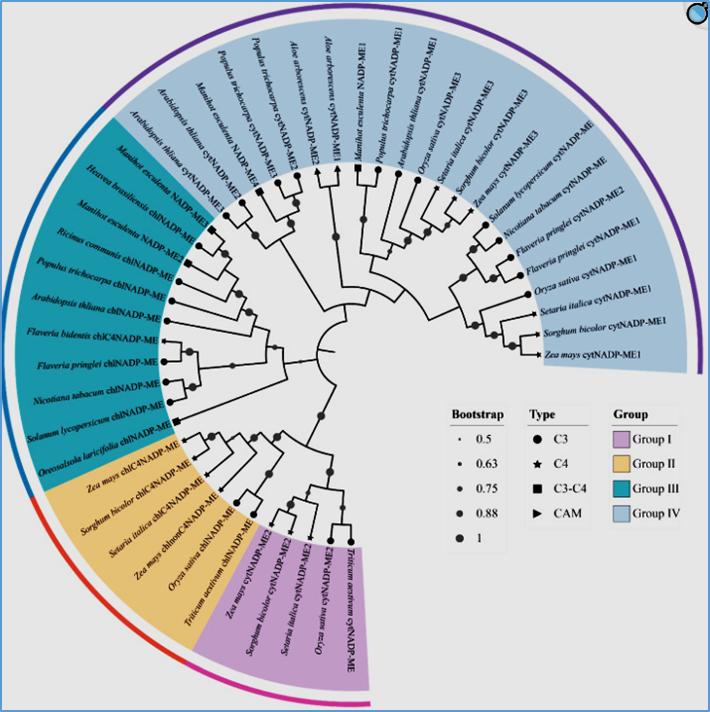
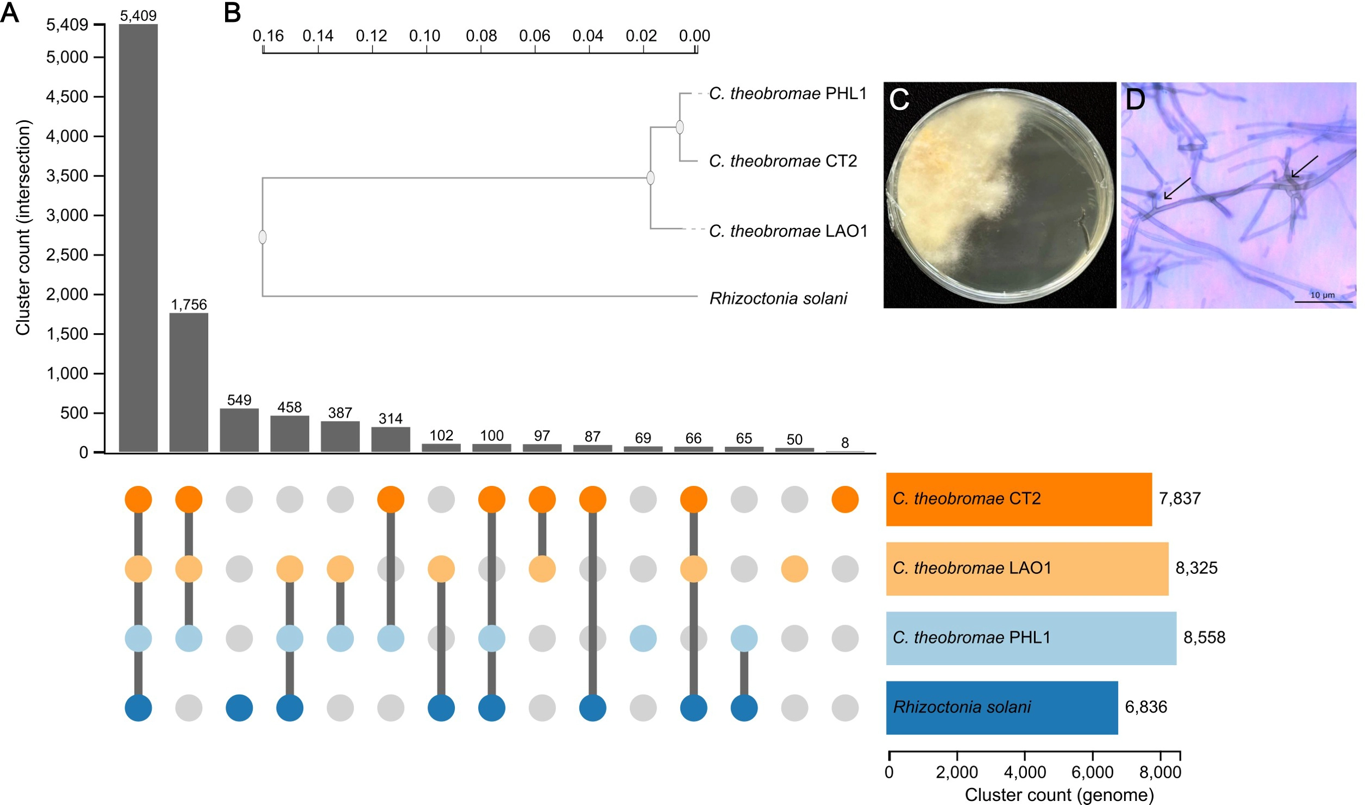
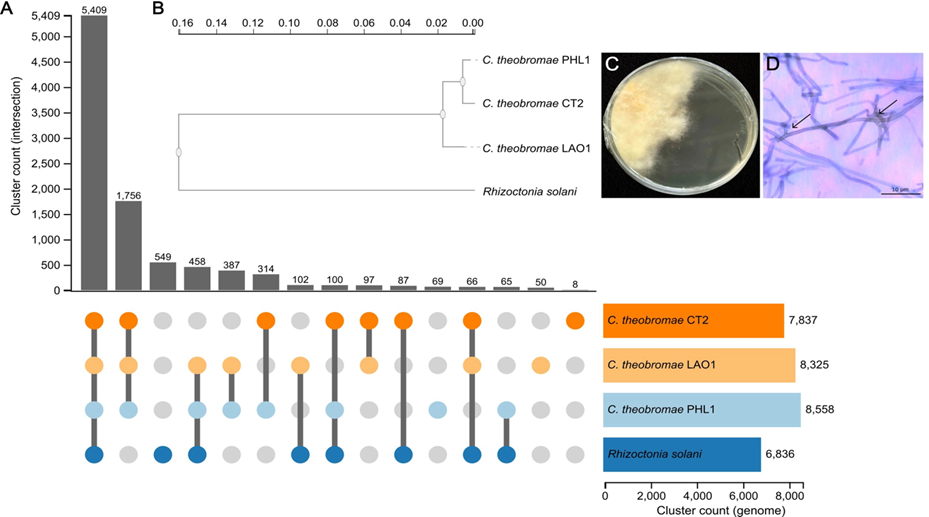
Giống sắn STB1 và KM444 trong mã hiệu sắn KM của VNCP
Link: https://vaas.vn/vi/giong/giong-san-stb1
1. Nguồn gốc
Giống
sắn STB1 ban đầu có tên là HL2004-32, là con lai của tổ hợp lai KM444
(thụ phấn tự do) do TS. Hoàng Kim lai tạo năm 2003 tại Trung tâm nghiên
cứu thực nghiệm Nông nghiệp Hưng Lộc. Từ năm 2004-2011, Trung tâm Nghiên
cứu và Phát triển cây có củ đã chọn lọc, đánh giá và khảo nghiệm dòng
sắn HL2004-32 ở các vùng sinh thái thuộc Trung du miền núi phía Bắc; Từ
năm 2012, Trung tâm nghiên cứu và phát triển cây có củ giao dòng sắn
HL2004-32 cho Viện Khoa học Kỹ thuật Nông nghiệp Bắc Trung Bộ đánh giá
và phát triển. Năm 2012, Viện KHKT Nông nghiệp Bắc Trung Bộ đã đổi tên
dòng sắn HL2004-32 thành giống sắn STB1 và tiến hành đánh giá giá trị
canh tác và sử dụng của giống sắn trên. Giống sắn STB1 đã được Bộ Nông
nghiệp và PTNT công nhận sản xuất thử giống cây trồng nông nghiệp theo
quyết định số 411/QĐ-BNN-TT vào ngày 16 tháng 2 năm 2017.
Năm 2021, Viện Khoa học Kỹ thuật Nông nghiệp Bắc Trung Bộ ban hành tiêu chuẩn cơ sở cho giống sắn STB1 và tự công bố lưu hành theo thông báo số 1187/TB-TT-CLT ngày 22 tháng 11 năm 2021 của Cục Trồng trọt.
2. Tác giả
TS. Phạm Văn Linh1, ThS. Phạm Duy Trình1, ThS. Nguyễn Quang Huy1, ThS. Trần Thị Duyên1, ThS. Nguyễn Đức Anh1, ThS. Lê Văn Quốc1, ThS. Lê Thị Thơm1, ThS. Đào Thị Minh Hiền1, ThS. Dương Thị Khánh Ly1, TS. Phạm Hùng Cương1, ThS. Lê Thị Thanh Thuỷ1, ThS. Nguyễn Tất Hoá1, ThS. Nguyễn Văn Phường1, ThS. Nguyễn Thị Huyền Trang1, TS. Hoàng Trọng Kim2.
Cơ quan công tác của tác giả: 1 Viện Khoa học Kỹ thuật Nông nghiệp Bắc Trung Bộ; 2 Trung tâm nghiên cứu và Phát triển cây có củ, Viện Cây lương thực và Cây thực phẩm.
3. Đặc điểm chính
Giống
STB1 có dạng cây thẳng, ít phân cành, chiều cao trung bình 285cm,
màu sắc thân xanh xám, màu sắc ngọn tím, vỏ củ màu
xám,..đường kính thân to, khả năng chống gãy đổ tốt, nhiễm nhẹ với một
số sâu bệnh hại chính, có khả năng thích ứng trên đất đỏ bazan, đất bạc
màu ven biển 2 tỉnh Hà Tĩnh và Nghệ An. Hàm lượng tinh bột của giống sắn
STB1 đạt 30,92 đến 33,82%, năng suất củ tươi của giống sắn STB1 đạt 38,7 – 52,3 tấn/ha.
4. Kỹ thuật trồng và chăm sóc
4.1. Chuẩn bị giống, đất trồng
Chuẩn
bị hom: Chọn những cây sắn khỏe mạnh, không bị nhiễm sâu bệnh, không bị
trầy xước, có đốt ngắn, ở những nương sắn tốt, không bị sâu bệnh, đã đủ
8 tháng tuổi trở lên để làm giống. Chọn lấy đoạn giữa thân, để chặt hom
làm giống. Dùng dao sắc chặt hom, mỗi hom dài 15 – 20 cm, có 3 – 5 mắt.
Khi chặt tránh làm dập nát hai đầu, tránh làm xây xước hom.
Xử lý hom giống trước khi trồng bằng cách nhúng vào các thuốc diệt nấm Ridomil hoặc Tilsuper 300EC để hạn chế sâu bệnh hại sắn.
Lưu ý: Các giống sắn đưa vào trồng phải đảm bảo sạch bệnh, đặc biệt chú ý đối với bệnh chổi rồng và khảm lá sắn.
Đối
với đất <15 độ, cày sâu và bừa kỹ để đất tơi xốp, thoáng khí và sạch
cỏ dại, nếu đất bằng thì nên lên luống để thoát nước. Đối với đất dốc
>15 độ, không cày bừa mà chỉ cần làm cỏ, bổ hốc và trồng sắn trực
tiếp theo đường đồng mức nhằm tránh xói mòn đất.
Giống
sắn STB1 trồng được trên nhiều loại đất khác nhau như: đất rừng mới
được khai thác, đất luân xen canh với các loại cây công nghiệp, cây thực
phẩm (cây họ đậu, lúa nước) và đất hoang hóa. Do nhu cầu để hình thành
và phát triển rễ củ, cây sắn cần đất tơi xốp thông thoáng. Đối với đất
trồng sắn trên các chân ruộng luân canh lúa nước thì sau khi nước rút và
thu hoạch lúa cần chuẩn bị đất sớm để xuống giống nhằm tranh thủ và tận
dụng được ẩm độ đất, gồm các khâu: xử lý cỏ dại, san lấp mặt bằng (nếu
đất bị úng cục bộ có thể vét mương hoặc rãnh thoát nước), cày hoặc phay
đất sớm và kéo liếp ngay sau khi nước rút.
4.2. Thời vụ
Thời
vụ trồng tốt nhất của giống sắn STB1 là vụ Xuân (10/01 – 20/03). Các
tỉnh Quảng Trị thì bố trí trồng sớm hơn từ 01/01 – 10/02 tránh khô hạn.
4.3. Kỹ thuật trồng
Có
ba phương pháp trồng hom sắn: Trồng hom nằm ngang trên những diện tích
đất tương đối bằng phẳng, ở những diện tích đất có mưa nhiều thoát nước
kém có thể kéo luống hoặc lên líp để trồng với các phương pháp hom xiên
hoặc hom đứng. Ngoài ra, nếu trồng vào vụ cuối mưa ẩm độ đất thấp thì
nên trồng hom đứng hoặc xiên.
4.4. Kỹ thuật bón phân
Tùy
theo các loại đất mà bón với các công thức khác nhau, có thể kết hợp
giữa bón phân vô cơ với phân hữu cơ như: phân chuồng, phân xanh và các
loại phân hữu cơ dạng lỏng.
Công thức bón NPK cho 1 ha sắn là: 90kg N + 60kg P2O5 + 90kg K2O + 15 tấn phân chuồng.
Thời
gian bón: Bón lót toàn bộ phân lân và phân hữu cơ. Bón thúc lần 1 vào
giai đoạn từ 25 - 30 ngày sau khi mọc mầm: ½ phân đạm + ½ phân kali, bón
thúc lần 2 vào giai đoạn sau khi mọc mầm từ 50 - 60 ngày: ½ phân đạm + ½
phân kali còn lại.
Thời điểm bón: Bón khi đất có đủ ẩm độ, tránh bón phân vào lúc trời nắng hoặc đang mưa lớn.
Phương
pháp và kỹ thuật bón: phân lân và phân hữu cơ bón lót khi cày bừa hoặc
bón theo hàng hay hốc trước khi trồng; phân đạm và phân kali bón theo
hốc (cuốc hốc cách gốc hoặc hom sắn 15cm rải đều phân xuống và lấp lại).
4.5. Kỹ thuật chăm sóc
Xới phá váng, làm sạch cỏ dại sau trồng từ 15-20 ngày.
Khi sắn có từ 5-7 lá (sau mọc mầm từ 30-40 ngày): làm sạch cỏ, xới đất, bón thúc lần 1 và vun nhẹ quanh gốc.
Khi sắn có từ 9-10 lá (sau mọc mầm từ 50-70 ngày): làm sạch cỏ, xới đất, bón thúc lần 2 và vun cao chống đổ.
4.6. Phòng trừ sâu bệnh
Một
số bệnh hại trên sắn: bệnh cháy lá do vi khuẩn, bệnh đốm lá, bệnh chổi
rồng. Biện pháp phòng trừ tốt nhất là sử dụng cây giống sạch bệnh, bón
phân cân đối, đầy đủ.
Một
số sâu hại trên sắn: mối, rệp sáp; trong đó mối là loại sâu hại chủ yếu
và quan trọng trên sắn. Mối gây hại chủ yếu ở giai đoạn mới trồng và
quá trình bảo quản. Để phòng trừ mối gây hại, sử dụng Diazan 10H từ 3 -
5kg/ha rải vào đất khi cày bừa hoặc theo hốc lúc trồng.
5. Thu hoạch và bảo quản
Thu
hoạch đúng thời điểm khi hàm lượng tinh bột trong củ đạt từ 27 - 30%,
hoặc khi cây đã rụng gần hết lá ngọn (còn lại khoảng 6 - 8 lá) và lá đã
chuyển từ màu xanh sang vàng nhạt. Có nhiều phương pháp thu hoạch khác
nhau: thu hoạch bằng cơ giới, các dụng cụ thủ công và nhổ trực tiếp bằng
tay. Thu hoạch đến đâu vận chuyển ngay đến các cơ sở chế biến, tránh để
lâu hoặc phơi nắng ngoài đồng làm giảm hàm lượng tinh bột trong củ. Đối
với trường hợp bán sắn lát, sắn thu hoạch đến đâu thì phải tiến hành
xắt lát rồi phơi khô tại ruộng. Sắn lát khô với ẩm độ từ 11 - 12% có thể
đem bán ngay hoặc bảo quản trong bao - kho chứa, cần xử lý các loại
thuốc xông hơi để phòng trừ côn trùng và mọt.
6. Địa phương đã áp dụng thành công
Các tỉnh Nghệ An và Quảng Trị.
7. Địa chỉ liên hệ giống
Viện Khoa học kỹ thuật nông nghiệp Bắc Trung Bộ,
Số 586 đường Nguyễn Trường Tộ, Tp Vinh, Nghệ An.
Điện thoại: 02383.514.625 / 0919.207.789.